
Role of Glucosinolates in Plant Stress Tolerance
P.S. Variyar, A. Banerjee, Jincy J. Akkarakaran and P. Suprasanna
Plant growth and development are affected by a variety of biotic and abiotic stresses. A multitude of responses are elicited during the adaptation of plants, which include activation of a defense system and a consequent enhanced production of secondary metabolites such as amino acids, sugars, indoles, phenolics, and glucosinolates. Glucosinolates are one of the major metabolites of family Brassicaceae biosynthesized from amino acids and responsible for the flavor and odor of Brassica vegetables. These compounds are induced after wounding and pathogen/insect attack as well as under abiotic stresses such as salt stress, UV radiation, and by plant signaling molecules. Following tissue damage, glucosinolates are hydrolyzed in vivo by endogenous enzymes, myrosinase, to unstable aglycones that further rearrange to a variety of products including isothiocyanates, thiocyanates, and nitriles. Glucosinolate levels and structure are also known to influence host plant suitability for the generalist and specialist herbivore. Levels of these compounds are affected under abiotic stresses including temperature and heavy metal stress. The concentration and type of glucosinolates and their hydrolysis are regulated at the genetic, environmental, and developmental levels. The regulation of glucosinolate metabolism at different levels and the diverse physiological function of their metabolites indicate a complex metabolic network. There is a paucity of information at the molecular level on the functional aspects such as signaling transduction pathways, control at transcriptional, translational, and post-translational levels, subcellular compartmentation, and interaction with many other metabolic pathways. Further knowledge on these and related aspects can aid in metabolic engineering of Brassica crops for stress tolerance and quality.
Keywords
Glucosinolates; abiotic stress; biotic stress; myrosinase; isothiocyanates
12.1 Introduction
Plants elicit multiple responses when exposed to a complex array of biotic (e.g., pathogen infection and herbivore feeding) and abiotic (e.g., nutrient levels and light conditions) stress factors. These stress factors induce signaling cascades that activate ion channels, kinases, production of reactive oxygen species (ROS), and accumulation of hormones such as salicylic acid (SA), ethylene (ET), jasmonic acid (JA) and abscisic acid (ABA) (Mittler, 2006; Jain, 2013). These signals eventually induce expression of specific subsets of defense genes that produce an overall defense response (Mittler et al., 2004; Mantri et al., 2011; Arbona et al., 2013). Activation of defense systems affects both the primary and secondary metabolism resulting in a substantial and significant variation in plant metabolome within and between species. Chemical defenses form a part of the plant’s inherent immune system. Plants produce a wide diversity of secondary metabolites that play a prominent role in defense against herbivores and pathogens. Some of them also act as defense against abiotic stress and in communication with its own species and with other organisms. Plant secondary chemistry is phenotypically plastic and varies in response to both biotic and abiotic factors. The major classes of secondary metabolites include the terpenoids and phenolics as well as the nitrogen (N)- and sulfur (S)-containing compounds synthesized primarily from amino acids.
Brassicaceae plants that include cruciferous vegetables such as cabbage, broccoli, cauliflower, kale, etc. are some of the most popular vegetables consumed the world over and considered to be a good source of bioactive phytochemicals. Sulfur-containing glucosides – glucosinolates (GSLs) – are one of the most important phytochemicals of Brassica vegetables responsible for their characteristic flavor and odor (Fahey et al., 2001; Martínez-Ballesta et al., 2013). Structurally, they are anions made up of thiohydroximates containing an S-linked β-glucopyranosyl residue and an O-linked sulfate residue with a variable amino acid-derived side chain. Unlike the other major classes of natural plant products, glucosinolates comprise a relatively small but diverse group of secondary metabolites that are generally limited to the species of the order Brassicales. These compounds represent a large chemical family that includes over 130 different compounds with varying structural subgroups (Clarke, 2010). They are largely responsible for the nutraceutical and pharmacological value of Brassica vegetables and have been implicated in defense against insects and pathogens and thus possess a bio-protective role. Efforts to improve specific quality attributes of plant foods, for example GSL, through breeding for quantitative food processing traits, are both promising and challenging (Hennig et al., 2013; Banerjee et al., 2014).
The level of glucosinolate metabolites has been shown to be induced upon root colonization by Trichoderma (Brotman et al., 2013). Following tissue damage, GSLs are hydrolyzed in vivo by endogenous enzymes, myrosinase, to unstable aglycones that further rearrange to a variety of products including isothiocyanates, thiocyanates, and nitriles, the nature of which depends upon the condition of hydrolysis and the structure of the GSLs (Halkier and Gershenzon, 2006). GSLs and their hydrolytic products have been documented to have significant antimicrobial and insecticidal activities (Aires et al., 2009). These compounds are known to accumulate in Brassica tissues after infestation by various pathogens restricting either the spread of fungal infection or inhibiting subsequent infections. Profound (and species-specific) temporal changes in GSLs have also been reported during insect herbivory (Yan and Chen, 2007). Glucosinolate structure and levels have been shown to influence host plant suitability for generalist and specialist herbivore and their levels were altered in response to herbivory (Halkier and Gershenzon, 2006).
The concentration and type of GSLs and their hydrolysis in plants has been shown to be regulated by genetic fluctuation, by environmental factors as well as by developmental cues (Martínez-Ballesta et al., 2013). Changes in total as well as different GSL subgroups have been reported in broccoli as a result of changes in salinity suggesting a role for these compounds in the leaf water response (Martínez-Ballesta et al., 2013). Induction of GSLs in broccoli mediated by ultraviolet-B (UV-B) was found to be associated with up-regulation of genes responsive to fungal and bacterial pathogens, thus demonstrating their role as stress alleviators (Mewis et al., 2012a). These compounds are induced in response to plant signaling molecules such as SA, JA, and methyl jasmonate (MeJA), the nature and extent of elicitation being dependent on the type of elicitors (Yan and Chen, 2007). Levels of these compounds are reported to be effected under temperature and heavy metal stress and also by post-harvest storage conditions (Yan and Chen, 2007). Thus, while selenium was found to affect the content of glucosinolates in a concentration-dependent manner, cadmium stress produced no change in glucosinolate production in B. rapa (Kim and Juvic, 2011; Jakovljević et al., 2013). Glucosinolate concentration also increased as a result of temperature stress showing seasonal variation in Brassica plants (Martínez-Ballesta et al., 2013).
Van Dam et al. (2009) summarized information on the GSL levels in both root and shoot in the same plant. The authors analyzed constitutive root and shoot glucosinolates of 29 plant species, which showed that roots have higher concentrations and a greater diversity of glucosinolates than shoots. Roots have significantly higher levels of the aromatic 2-phenylethyl glucosinolate, possibly related to the greater effectiveness and toxicity of its hydrolysis products in soil. It was also seen that in shoots, the most dominant indole glucosinolate is indol-3-ylglucosinolate, whereas in roots, its methoxy derivatives are dominated. The regulation of GSL metabolism at different levels and the diverse physiological function of their metabolites indicate a complex metabolic network. Studies at the genetic level have shown that GSL metabolism interacts with cellular signaling and metabolic pathways and is regulated at different levels. The extent of GSL hydrolysis that effects interaction with microbes and herbivores is also known to be controlled at the genetic level (Sønderby et al., 2010). There is, however, a lack of understanding at the molecular level on the functional aspects such as signaling transduction pathways, control at transcriptional, translational and post-translational levels, subcellular compartmentation, and interaction with many other metabolic pathways. Further studies are needed to understand the sophisticated signaling network that connects environmental factors with GSL metabolism. Knowledge on these and related aspects can aid in metabolic engineering of Brassica crops for better quality, nutrition, and disease resistance. Considerable interest in optimizing GSL content and composition for plant protection and human health has made GSLs a dynamic area in plant metabolomic research.
12.2 Glucosinolate structure, isolation, and analysis
The first general structure of GSL was proposed in 1897 by Gadamer (Fahey et al., 2001), which proposed that the side chain is linked to the nitrogen rather than to the carbon atom (Figure 12.1). Several approaches have been proposed for classification of GSLs into subgroups. The most common approach is based on the nature of the biosynthetic precursor amino acids. Another approach classifies them into aliphatic, aromatic, and indolic, or aliphatic, benzenic, and indolic derivatives. They are also grouped based on their tendency for forming specific breakdown products. Specific nomenclature of individual GSLs is generally based on naming the entire anionic structure (the central carbon (C) as well as the connected, substituted S and N) as glucosinolate and adding the systematic name of the side chain as a radical.
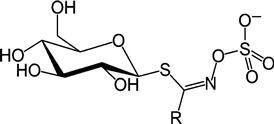
Glucosinolates are known to be regulated both developmentally and environmentally in various organs and tissues depending on the type of biotic and abiotic stresses. Depending on the developmental stage, tissue, and photoperiod, the distribution pattern of GSLs differs between species and ecotypes as well as between and within individual plants (Table 12.1; Martínez-Ballesta et al., 2013). Tissue-level glucosinolate accumulation has a major influence on its hydrolysis to bioactive products. Vascular tissue has been found to be the site of glucosinolate biosynthesis with endoplasmic reticulum as the subcellular location of GSL biosynthetic enzymes. Plants containing GSLs always possess a thioglucosidase called myrosinase that catalyzes the degradation of GSL substrates when plant tissue is disrupted as a result of wounding or insect and pathogen attack (Sønderby et al., 2010). The majority of the myrosinase enzymes act on multiple GSL substrates, although some of them have high specificity. Myrosinases are localized in specific cells named myrosin cells or myrosin idioblasts that contain protein-rich vacuolar-type structures termed myrosin grains. Glucosinolates, however, have been reported to be localized in vacuoles in non-specific cells together with ascorbic acid, which modulates myrosinase activity.
Table 12.1
Glucosinolates Found in Different Food Sources
| Trivial Name | R Side Chain | Food Source |
| Glucocapparin | Methyl | Capers |
| Glucolepidin | Ethyl | Radish |
| – | Propyl | Cabbage |
| Glucoputranjivin | Isopropyl | Radish |
| Sinigrin | 2-Propenyl | Cabbage |
| Glucoiberin | 3-Methylsulfinylpropyl | Cabbage |
| Glucoibervirin | 3-Methylthiopropyl | Cabbage |
| Glucocheirolin | 3-Methylsulfonylpropyl | Cow’s milk |
| Glucocapparisflexuosain | Butyl | Cabbage |
| Gluconapin | 3-Butenyl | Cabbage |
| Progoitrin | (2R)-2-Hydroxy-3-butenyl | Cabbage |
| Epiprogoitrin | (2S)-2-Hydroxy-3-butenyl | Sea kale |
| Glucoerucin | 4-Methylthiobutyl | Cabbage |
| Glucoraphanin | 4-Methylsulfinylbutyl | Broccoli |
| Glucoerysolin | 4-Methylsulfonylbutyl | Cabbage |
| Dehydroerucin | 4-Methylthiobut-3-enyl | Daikon’s radish |
| Glucoraphenin | 4-Methylsulfinylbut-3-enyl | Radish |
| Glucobrassicanapin | 4-Pentenyl | Chinese cabbage |
| Glucoberteroin | 5-Methylthiopentyl | Cabbage |
| Glucoalyssin | 5-Methylsulfinylpentyl | Rocket |
| Gluconapoleiferin | 2-Hydroxy-pent-4-enyl | Swede |
| Glucosiberin | 7-Methylsulfinylheptyl | Watercress |
| Glucohirsutin | 8-Methylsulfinyloctyl | Watercress |
| 4-Hydroxyglucobrassicin | 4-Hydroxy-3-indolylmethyl | Cabbage |
| Glucobrassicin | 3-Indolylmethyl | Cabbage |
| 4-Methoxyglucobrassicin | 4-Methoxy-3-indolylmethyl | Cabbage |
| Neoglucobrassicin | N-Methoxy-3-indolylmethyl | Cabbage |
| Glucotropaeolin | Benzyl | Cabbage |
| Glucosinalbin | p-Hydroxybenzyl | Mustard |
| Gluconasturtiin | 2-Phenylethyl | Cabbage |
| Glucobarbarin | (2S)-2-Hydroxy-2-phenylethyl | Land cress |
| Glucosibarin | (2R)-2-Hydroxy-2-phenylethyl | White mustard |
Substantial degradation of GSLs by myrosinase enzymes when extracted with cold organic solvent necessitates the use of hot aqueous alcohols such as methanol:water (70:30) for their isolation from plant materials (Clarke, 2010). This process denatures the enzyme and prevents hydrolysis of these compounds thereby facilitating their quantitative isolation. A prior separation into groups normally precedes their identification and quantification by instrumental methods. The presence of sulfate groups facilitates binding of these compounds to an anion exchange column and thus allows separation of either the intact GSLs or “desulfo” derivatives after enzymatic desulfation (Clarke, 2010). Direct analysis of volatile isothiocyanates and nitriles produced from GSLs by gas chromatography-mass spectrometry (GC-MS) can also provide proof of the presence of corresponding GSL in intact plants. Use of high-pressure liquid chromatography-mass spectrometry (HPLC-MSn) for detection of intact GSLs in crude extracts is currently a powerful routine method comparable in specificity with the classical methods of identification (Clarke, 2010). Capillary electrophoresis for simultaneous quantification of GSLs and their hydrolysis products has also been reported (Clarke, 2010). Use of modern MSn equipment with ion traps allows for highly sophisticated analysis of side chain structures and validation of elucidated GSL structures. Even with highly sophisticated MS-based detection methods, comparison of chromatographic retention time with authentic standard and one additional characteristic property such as retention time in a different chromatographic system, a characteristic UV spectrum, a mass spectrum, or nuclear magnetic resonance (NMR) data is a must to suggest a tentative identification of a given GSL.
12.3 Biosynthesis of glucosinolates
Biosynthesis of GSLs involves three independent stages, namely: (1) chain elongation of selected precursor amino acids (mainly methionine) by addition of methylene groups; (2) formation of core glucosinolate structure by reconfiguration of the amino acid moiety; and (3) secondary modification of the amino acid side chain by hydroxylations, methylations, oxidations, or desaturations. While the construction of core anionic structure from amino acids involves a number of common steps, a number of diverse steps are involved in formation of side chain and other diversifications. Aliphatic GSLs are derived from alanine, leucine, isoleucine, valine, and methionine, while benzenic GSLs are formed from phenylalanine and tryptophan and indolic GSLs from tryptophan (Sønderby et al., 2010).
Synthesis of the core GSL structure is achieved in five steps (Figure 12.2). The first step involves oxidation of precursor amino acids to aldoximes by side chain-specific cytochrome P450 monooxygenase of the CYP79 family. Further oxidation by cytochrome P450 of the CYP83 family leads to aci-nitro compounds or nitrile oxides. The nitro compounds formed are strong electrophiles that react spontaneously with thiols to form S-alkylthiohydroximate conjugates that further undergo cleavage into unstable thiohydroximates, pyruvate, and ammonia by the action of a C–S lyase. Glucosyl transferase catalyses thiohydroximate-specific S-glycosylation. The final step is the 3′-phosphoadenosine 5′-phosphosulfate-dependent sulfation of desulfoglucosinolates (Sønderby et al., 2010).

The amino acid elongation is similar to the valine-to-leucine conversion and involves five steps which include initial and final transamination, acetyl-CoA condensation, isomerization, and oxidative decarboxylation. Methylthioalkylmalate (MAM) synthases that catalyze the condensation reaction have been characterized in Arabidopsis and Eruca sativa. Methionine side chain elongation occurs in the chloroplast and elongated α-keto acid can either be transaminated and enter the core GSL pathway or undergo additional elongation steps with insertion of up to nine methylene units. The variation in side chain length of methionine-derived GSL is controlled by three partially redundant MAM genes (Sønderby et al., 2010).
Secondary modification of the side chains involving various types of oxidations, eliminations, alkylations, and esterifications is generally considered as the final stage in GSL synthesis. An extensive natural variation of aliphatic glucosinolates has been noted in Arabidopsis with two α-ketogluterate-dependent dioxygenases controlling the production of alkenyl and hydroxyalkyl GSLs (Kliebenstein et al., 2001).
An interdependent metabolic control of aliphatic and indolyl GSL branches has been proposed indicating a homeostatic control of GSL synthesis. This is achieved by a reciprocal negative feedback regulation between both the branches using intermediates or end products of glucosinolate biosynthesis as inhibitors. Limited NADPH supply has also been proposed for the interdependence of the two pathways wherein inhibition of one branch would lead to increased NADPH availability for the other. In addition, side chain elongation can lead to extra yield of NADH that can be converted to NADPH via the malate dehydrogenase and maleic enzyme reactions. Thus, side chain elongations can provide NADPH independently of the pentose-phosphate pathway thereby increasing GSL production. Considerable variation is thus noted in the total as well as individual GSL content of methionine-derived and indolyl GSL in leaves and seeds, respectively (Grubb and Abel, 2006).
12.4 Role of glucosinolates in stress alleviation
Loss of cellular integrity as a consequence of stress induced by wounding, insect, or pathogen attack leads to hydrolysis of GSLs by the enzyme myrosinase. GSLs and their hydrolytic products are frequently investigated for their role as a plant defense system against insects, herbivores, and certain microbial pathogens. It has been shown that infection with fungal pathogen can induce local synthesis of myrosinase and the possibility of such a mechanism under other stress response is also proposed. Environmental factors influence secondary metabolism as plants under stress produce more secondary metabolites, more so as the growth is often limited more than in photosynthesis, and carbon fixation is predominantly invested to secondary metabolite production (Endara and Coley, 2011). It has been very well reported that environmental factors, such as light (Engelen-Eigles et al., 2006), temperature (Velasco et al., 2007), salinity (Qasim et al., 2003; López-Berenguer et al., 2009), water (Champolivier and Merrien, 1996; Rask et al., 2000), CO2 (Schonhof et al., 2007a), and drought (Radovich et al., 2005) may affect glucosinolate levels (Table 12.2).
Table 12.2
Impact of Abiotic Stress on Glucosinolate Accumulation in Different Brassica Plant Species
| Plant Species | Glucosinolate Content | Stress | Treatment Condition |
| Brassica oleracea L. var. italic | Increase | Salinity | NaCl (40, 80 mM), during 2 weeks |
| Brassica rapa L. | Increase | NaCl (20, 40, 60 mM), during 5 days | |
| Brassica campestris L. ssp. chinensis var. communis | NaCl (50 and 100 mM for 2 weeks) | ||
| Brassica oleracea L. var. capitata | Increase | Drought | Severe stress 2 weeks |
| Brassica oleracea L. var. italica | Increase | Severe stress 2 weeks | |
| Brassica napus L. | Increase | Severe stress more than 1 week | |
| Brassica rapa ssp. rapifera L. | Increase | Mild stress—25% of available water | |
| Brassica carinata L. | Increase/no effect | Mild and severe stress (40, 23, 17 and 15% of available water) | |
| Brassica oleracea L. var. gemmifera | No effect | Mild stress (30% of available water) | |
| Brassica napus L. | No effect | Mild stress | |
| Brassica oleracea L. | Decrease | Mild and severe stress (40–45% of available water) | |
| Arabidopsis thaliana L. | Decrease | Severe stress | |
| Arabidopsis thaliana (L.) | Decrease | Mild stress (50% of available water) | |
| Arabidopsis thaliana (L.) | Decrease | Water logging (200% of available water) | |
| Brassica rapa L. | Increase | Temperature | Elevated temperature (21–34°C) |
| Brassica rapa L. | Decrease | Low–medium temperature (15–27°C) | |
| Brassica oleracea L. | Increase | Elevated temperature (32°C) | |
| Brassica oleracea L. | Decrease during day/increase during night | Light cycling | 14 h/10 h day/night* |
| Arabidopsis thaliana L. | Increase upon light/decrease upon darkness | 16 h/8 h d/n or continuous darkness | |
| Brassica oleracea L. var. italica | Increase upon light | 16 h/8 h d/n or continuous darkness | |
| Arabidopsis thaliana | Slight increase | UV-B radiation | 1.55 Wm−2 |
| Brassica oleracea L. var. italica | Increase | Up to 0.9 kJm−2 d−1 | |
| Brassica oleracea L. var. italica | Increase | Nutrient availability | N-limitation (1 gr N pot−1) |
| Brassica rapa ssp. rapifera L | Increase | S-supply (60 kg S ha−1) | |
| Brassica oleracea L. var. italica | No effect | S-supply (150 kg/ha) | |
| Brassica oleracea L. capitata | Increase | S-supply (110 kg S ha−1) | |
| Brassica napus | Increase | S-supply (100 kg S ha−1) | |
| Tropaeolum majus | Increase | S-supply (8.3 mM SO42−) | |
| Brassica oleracea L. var. italica | No effect | S-limitation (15 kg/ha) | |
| Arabidopsis thaliana L. | Increase | K-deficiency (lack KNO3 for 2 weeks) | |
| Brassica rapa L. | Decrease | K-deficiency (lack of nutrient solution for 5 days) | |
| Brassica oleracea L. var. italica | Increase | Se-supply (5.2 mM Na2 SeO4) | |
| Brassica oleracea L. var. italica | B-deficiency (9–12 μg gr DW−1) | ||
| Cabbage and kale | Increase | Cadmium | Cd (5 and 10 mg Cd kg−1 soil) |
| Thlaspi caerulescens | Increase |

Source: Modified after Martínez-Ballesta et al., 2013.
As Brassica crops contain high amounts of sulfur-containing amino acids and glucosinolates, glucosinolate metabolism and the effects of sulfur and nitrogen nutrition have been studied (Schnug et al., 1993; Krumbein et al., 2002; Salac et al., 2006; Schonhof et al., 2007b). It is evident that when broccoli plants were supplied with low sulfur or nitrogen, a decrease in glucosinolates was noted, whereas total glucosinolate levels were elevated at sufficient nitrogen supply or high sulfur levels, and were lower at low sulfur supply with an optimal nitrogen supply (Aires et al., 2006; Schonhof et al., 2007a). Similarly, glucosinolate levels in turnip were found to be strongly regulated by nitrogen and sulfur application (Kim et al., 2002). In field experiments, nitrogen and sulfur supply showed a clear influence on individual glucosinolates as it may favor the hydroxylation step converting but-3-enyl glucosinolate to 2-hydroxybut-3-enyl glucosinolate. Compared to indole glucosinolates, aliphatic glucosinolates show a greater sensitivity to sulfur deficiency probably because they are synthesized from methionine (Zhao et al., 1994). Some B. napus cultivars with reduced contents of aliphatic glucosinolates were more sensitive to sulfur deficiency (Schnug, 1990), which suggests a role of aliphatic glucosinolates in the survival strategy to mineral stress. Sulfur fertilisation leads to increases in glucosinolate content in most cases. Increases of over 10-fold have sometimes been reported. For example, the benzyl glucosinolate content of Tropaeolum majus was increased over 50-fold by fertilising a particular cultivar with 8.3mM sulfate (Matallana et al., 2006).
12.4.1 Biotic stress
During their lifetime, plants have to deal with a variety of environmental stresses including biotic stresses such as those from microbial pathogens and herbivores. As plants are not in a position to move from their unfavorable environment, they have evolved a broad range of defense mechanisms. The role of GSLs in combating biotic stress has been well recognized. GSLs exhibit growth inhibition or feeding deterrence to a wide range of general herbivores such as birds, slugs, and generalist insects (Rask et al., 2000; Barth and Jander, 2006). Plants respond to herbivore or insect damage by accumulating higher GSL levels and thus increase their resistance to such biotic stresses. Glucosinolates, the characteristic secondary compounds of Brassicaceae, as well as proteinase inhibitors, remained unaffected by UV in all plants, demonstrating independent regulation pathways for different metabolites (Kuhlmann and Müller, 2009a,b). Mewis et al. (2012b), however, demonstrated an increase in aliphatic GSLs in Arabidopsis thaliana when fed by phloem-feeding aphids, the green peach aphid (Myzus persicae), cabbage aphid (Brevicoryne brassicae), and generalist caterpillar species Spodoptera exigua. Interestingly, the content of indole GSLs were found to be unchanged. GSL levels have been demonstrated to reduce damage by generalist herbivores. Volatiles produced by GSLs can also provide indirect protection to plants by attracting natural enemies of herbivores such as parasitoids. Several reports exist on the toxicity of GSL hydrolysis products to bacteria and fungi (Mayton et al., 1996; Brader et al., 2001). Pedras and Sorensen (1998) demonstrated an inhibitory action by various isothiocyanates derived from GSLs on germination and growth of a fungal pathogen Leptosphaeria maculans. Aromatic isothiocyanates were found to be more toxic than aliphatic isothiocyanates and the fungal toxicity of the latter decreased with increase in side chain length. In a study on the antimicrobial effect of crude extracts from Arabidopsis, Tierens et al. (2001) identified 4-methylsulfonyl butyl isothiocyanate as the major active compound with a broad spectrum of antimicrobial activity. Thus, the possible protective role of GSL-derived isothiocyanate against pathogens was demonstrated. Investigation of the level of GSLs in different Brassica cultivars by several workers indicated changes in GSL pattern when inoculated by fungal pathogens. These changes were mostly due to increase of indole and aromatic GSLs, although increase of aliphatic GSLs was also noted.
12.4.2 Abiotic stress
All abiotic stresses are important environmental factors that reduce plant growth and yield. Plants respond and adapt to these stresses in order to survive. Signaling pathways are induced in response to environmental stresses. Several signaling molecules have been identified in plant defense responses. These include JA, SA, and ET, which have been demonstrated to operate independently and/or synergistically in different signal transduction pathways. JA and SA have been shown to be involved in the induction of different GSLs (Yan and Chen, 2007). Different signal transduction pathways activate specific biosynthetic and secondary modifying enzymes, leading to altered levels of specific GSLs. The induction of GSLs by several defense pathways strongly indicates that these compounds play a role in plant defense.
Salt stress is a major abiotic stress reducing the productivity of crops in many areas of the world. Salinity affects the water balance resulting in osmotic damage. Osmotic adjustment is a plant adaptation mechanism used to maintain water balance in plants. In their studies on the effect of salinity stress on GSL content, Keling and Zhujun (2010) found a considerable influence of NaCl stress on the GSL content and composition in pakchoi (Brassica campestris L. ssp. chinensis var. communis) shoots. At 50 mM NaCl, the contents of total GSLs as well as aliphatic and indole GSL significantly increased. A significant increase in indole GSLs and a decrease in aromatic GSL (gluconasturtiin) were, however, noted at 100 mM NaCl. Glucoalyssin, gluconapin, glucobrassicin, and neglucobrassicin were significantly enhanced at 50 mM NaCl, while only the content of gluconapin and glucobrassicin increased at 100 mM NaCl.
Drought stress resulted in considerably elevated leaf GSL content of Brassica carinata varieties with the magnitude of GSL concentration varying with the stage of development and intensity of the drought stress (Schreiner et al., 2009a). Increase in leaf GSL concentrations correlated with relative water content with reduced water content leading to higher leaf GSL concentration. Brassica oleracea L., plants grown for two weeks under drought stress showed decreased levels of indolyl GS when compared to well-watered plants, while water logging conditions resulted in slight increases within the GS contents (Khan et al., 2011). Imbalance in sulfur to nitrogen ratio may result in the alteration of nutrient uptake due to water deficit resulting in the accumulation of GSLs as sulfur sink. Further, stresses such as low water availability change the hormonal distribution of plants leading to a cascade of signal transduction pathways that result in the expression of stress-responsive genes. Particularly, stress hormones like ABA, JA, ethylene, and SA that play a very important role in biotic and abiotic stress resistance are known to increase the concentrations of GSLs (Yan and Chen, 2007).
UV-B radiation acts as an environmental stress and triggers various responses in plants. These include changes in growth, development, morphology, and physiological aspects. In recent years, some researchers have reported the effect of UV-B on GSL metabolism. Microarray data have shown that the genes related to the biosynthesis of flavonoids, glucosinolates, and terpenoids were differently expressed after UV-B radiation. A study on the effect of UV radiation on Tropaeolum majus demonstrated that appropriate UV-B dosage could increase the glucotropaeolin concentration (Schreiner et al., 2009b). Wang et al. (2011) showed that UV-B radiation induced the production of GSLs. Continuous UV-B exposure for 12 h, however, inhibited the expression of glucosinolate metabolism-related genes resulting in a significant decline in glucosinolate content, particularly that of indolic glucosinolates. In another study on UV-B-mediated induction of GSLs, Mewis et al. (2012c) reported the induction of of 4-methylsulfinyl butyl GSL and 4-methoxy-indol-3-ylmethyl GSL in sprouts of Brassica oleracea var. italica (broccoli). Accumulation of defensive GSLs was accompanied by increased expression of genes associated with salicylate and JA signaling defense pathways and up-regulation of genes responsive to fungal and bacterial pathogens. Enhanced GSL formation had a negative effect on the growth of aphid Myzuz persicae and attack by caterpillar Pieris brassicae. Levels of these compounds are also reported to be effected under temperature stress. The TU8 mutant of Arabidopsis deficient in glucosinolate metabolism and pathogen-induced auxin accumulation showed less tolerance to elevated temperatures than wild-type plants (Ludwig-Müller et al., 2000). Seasonal variation in the concentration of aliphatic, aromatic, and indole GSLs was noted in different varieties of Brassica oleraceae (Cartea et al., 2008). Similar effects with increase in aliphatic glucosinolates (particularly glucoraphanin) were observed in broccoli kept at daily mean temperatures between 7 and 13°C (mean radiation of 10–13 mol m−2 day−1) (Schonhof et al., 2007c).
In the authors’ laboratory (Banerjee et al., 2014), the cabbage leaves subjected to gamma radiation stress were found to have an enhanced sinigrin content. No effect of myrosinase activity was, however, noted, thus providing high retention of glucosinolates and facilitating improved release of these nutraceutically significant compounds during mastication of the vegetable. Thus, exposure to such abiotic stress was demonstrated to provide improved benefit in terms of enhancing intake of potentially important health protective and promoting compounds in Brassica vegetables (Banerjee et al., 2014).
Heavy metal stress also can lead to changes in GSL content. While selenium was found to affect the content of GSLs in a concentration-dependent manner, cadmium stress produced no change in GSL production in B. rapa (Kim and Juvic, 2011; Jakovljević et al., 2013). GSL concentration also increased as a result of temperature stress showing seasonal variation in Brassica plants. In Thlaspi caerulescens, a metal hyperaccumulator with a high requirement of zinc, GSL levels (particularly sinalbin) increased in roots but decreased in leaves and shoots. Zinc had a clearly distinctive effect on the specific group of indolyl GSLs in T. caerulescens with a drastic reduction in both roots. Post-harvest storage conditions of Brassica vegetables are also known to influence GSL and related isothiocyanate content. Content of these compounds was found to decrease in vegetables such as broccoli, brussel sprouts, cauliflower, and green cabbage when stored in a domestic refrigerator (4–8°C) for 7 days unlike when stored at ambient temperature (Song and Thornalley, 2007). Storage of vegetables at very low temperature (–85°C) can result in freeze–thaw fracture of plant cells leading to significant loss of GSLs as a consequence of their conversion to isothiocyanates during thawing (Song and Thornalley, 2007). Tamara et al. (2013) found that GSLs in leaves and root could be more involved in ameliorating S deficiency rather than plant defense in the short term in cadmium (Cd) stress; however, total GSL levels in the stem during the long term could serve as a GSL storage organ implying possible roles of GSL in Cd stress.
12.5 Genes involved in glucosinolate biosynthesis
The main genetic pathway of glucosinolate biosynthesis has been identified in Arabidopsis using genetic and biochemical approaches. Several enzymes and transcription factors involved in the GSL biosynthesis have been studied in the model plant, Arabidopsis, and in a few other Brassica crop species (Baskar et al., 2012). Figure 12.3 presents the genetic machinery involved in different aspects of GSL sysnthesis. Six MYB factors, namely, MYB28, MYB29, MYB76, MYB34, MYB51, and MYB122, have been found to be transcriptional regulators in the biosynthesis of glucosinolate in Arabidopsis. While MYB28, MYB29, and MYB76 specifically transactivate genes related to aliphatic glucosinolate biosynthetic pathway (MAM3, CYP79F1, and CYP83A1) (Gigolashvili et al., 2007b, 2008), MYB34, MYB51, and MYB122 are regulators of the indolic glucosinolate biosynthetic pathway (TSB1, CYP79B2, and CYP79B3) (Celenza et al., 2005; Gigolashvili et al., 2007a). Wang et al. (2011) used the comparative genomic analysis method of Arabidopsis thaliana and Brassica rapa and identified 102 putative genes in B. rapa as the orthologues of 52 Arabidopsis glucosinolate genes. The glucosinolate genes in B. rapa and A. thaliana shared 59–91% nucleotide sequence identity. Microarray experiments have also shown that CYP79B2, an important gene involved in the biosynthesis of indolic glucosinolates (Chen and Andreasson, 2001), is downregulated by brassinosteroids (Goda et al., 2002). Both MYB34 and MYB51, which encode transcriptional factors of indolic glucosinolate biosynthesis, contain a BZR1 binding site in their promoters (Sun et al., 2010). Further, Guo et al. (2012) investigated the role of brassinosteroids in glucosinolate biosynthesis in Arabidopsis using mutants and transgenic plants. Zang et al. (2009) identified glucosinolate synthesis genes in Brassica rapa on the basis of cDNA and BAC libraries with about 21.5% of the genes as partial CDS sequences and many BrGS genes anchored only on the BAC, rather than on chromosomes. The authors also identified glucosinolate biosynthetic genes by comparative genomic analysis between B. rapa and A. thaliana. Augustine et al. (2013) analyzed four MYB28 genes that are differentially expressed and regulated in both a tissue- and cell-specific manner in controlling aliphatic glucosinolate biosynthesis in B. juncea.

Several myrosinase genes from Sinapis alba, Brassica napus, and Arabidopsis thaliana have been isolated and characterized indicating that myrosinases are encoded by a multigene family consisting of three subgroups (Xu et al., 2004). Myrosinase in the Brassica family is encoded by a gene family, which consists of three subfamilies, namely, MA (Myr1), MB (Myr2), and MC (Baskar et al., 2012). Several myrosinase-associated proteins, such as epithiospecifier modifier 1 (ESM1), ESP, and MVP1, have been identified in Arabidopsis, which are mainly involved in the generation of diversified GSL metabolic products (Baskar et al., 2012).
12.6 Gene expression profiling in response to environmental cues
Plant glucosinolate metabolism is responsive to many environmental factors. Generally, glucosinolate degradation products serve as defense compounds against pathogens and generalist herbivores, and as attractants to glucosinolate-adapted specialists (Rask et al., 2000; Barth and Jander, 2006). Several glucosinolate hydrolysis products have been reported to display toxicity to fungi and bacteria (Mayton et al., 1996; Brader et al., 2001). Glucosinolate levels in oilseed rape were positively correlated with resistance to pathogens (Li et al., 1999) with some exceptions (Giamoustaris and Mithen, 1997). The best in vivo evidence for the defense role of glucosinolates came from an MAM1 mutant study, where decreased glucosinolate levels in Arabidopsis caused susceptibility to Fusarium oxysporum (Tierens et al., 2001). Pathogen infection can also change glucosinolate profiles. When Brassica plants were infected by Leptosphaeria maculans, glucosinolate levels were induced, but myrosinase levels were not affected (Siemens and Mitchell-Olds, 1998). JA and SA signaling pathways may be involved in the regulation of glucosinolate levels (Li et al., 2006). Currently, there is more literature on plant interactions with insect herbivores. When glucosinolate levels increased in B. napus and Sinapis alba, feeding by generalist insects decreased significantly, while feeding by specialist insects (e.g., Pieris rapae) greatly increased and caused severe damage (Giamoustaris and Mithen, 1995). The damage led to a systemic increase in indole glucosinolate and often in total glucosinolate levels. For example, when seedlings of oilseed rape and mustard were attacked by Xea beetles, there was a tremendous increase in the concentration of indole glucosinolates, but no significant changes in aliphatic glucosinolates (Bodnaryk, 1992; Bartlet et al., 1999). In one case when feeding with turnip root fly, the concentrations of aliphatic glucosinolates actually dropped (Hopkins et al., 1998).
Mewis et al. (2006) analyzed glucosinolate accumulation levels and gene expression of glucosinolate biosynthetic genes in response to feeding by four herbivores in Arabidopsis thaliana (L.) wild-type (Columbia) and mutant lines that were affected in defense signaling. Herbivory on wild-type plants led to increased aliphatic glucosinolate content for three of four herbivores tested, namely, Myzus perscae (Sulzer), Brevicoryne brassicae (L.), and Spodoptera exigua Hübner. The lepidopteran Pieris rapae L. did not affect the levels of aliphatic glucosinolate in the wild type, except for an increase in indole glucosinolates. Increased expression of genes of aliphatic glucosinolate biosynthesis was observed after feeding by all species, while mutations in jasmonate (coi1), salicylate (npr1), and ethylene signaling process (etr1) showed varied gene expression, glucosinolate profile, and insect performance compared to wild type. As against in wild type, the gene transcripts of aliphatic glucosinolate biosynthesis did not generally increase in the mutants. Both glucosinolate content and gene expression data indicated that salicylate and ethylene signaling repress some jasmonate-mediated responses to herbivory. This possibly indicates that all three modes of signaling processes are involved in responses to herbivores.
Plant interactions with the environment influences glucosinolate metabolism and is constantly regulated by different environmental factors including UV-B radiation. Wang et al. (2011) studied the glucosinolate content and expression of glucosinolate metabolism-related genes in response to enhanced UV-B radiation (1.55 W • m−2) and the succeeding dark recovery process in Arabidopsis thaliana rosette leaves. Induction of glucosinolates was observed in the 1 h of enhanced UV-B radiation exposure, whereas, after continuous exposure for 12 h, the expression of glucosinolate metabolism-related genes was significantly inhibited and the glucosinolate content was declined, especially that of indolic glucosinolates. Upon exposure to darkness for 12 h for partially recovery, both glucosinolate gene expression and the content returned to the control levels. The results of Wang et al. (2011) showed up-regulation of some genes (MYB51, OBP2, MYB76, SOT16, and TGG1) in the initial 1 h of UV-B exposure. These genes were also induced by JA and wounding (Skirycz et al., 2006; Dombrecht et al., 2007; Staswick, 2008). The transcription factors (MYB51, OBP2, and MYB76) can positively mediate glucosinolate biosynthesis (Skirycz et al., 2006; Dombrecht et al., 2007; Gigolashvili et al., 2008). MYB51 and MYB76 specifically activate indolic and aliphatic glucosinolate biosynthesis, respectively, which might be responsible for the temporary and initiative increase of 4MSOB and I3M. The 3 h of treatment led to down-regulation of MYB28 and MYB29 (the genes encoding the transcription factors that regulate aliphatic glucosinolate biosynthesis) and MYB34 (regulating indolic glucosinolate biosynthesis), and structural genes (CYP79F1, CYP83B1, MAM1, CYP79B2, and CYP79B3) encoding enzymes catalyze the synthesis of aliphatic glucosinolates. Subsequent exposure led to decline in the expression of the majority of genes and the glucosinolate contents. Microarray analysis experiments with UV-B exposed Tropaeolum majus also indicated differential expression of genes related to the biosynthesis of flavonoids, glucosinolates, and terpenoids (Hectors et al., 2007).
Schweizer et al. (2013) showed that a triple mutant for MYC2, MYC3, and MYC4, three basic helix–loop–helix transcription factors that are known to additively control jasmonate-related defense responses, was shown to have a highly reduced expression of GSL biosynthesis genes. The myc2 myc3 myc4 (myc234) triple mutant was almost completely devoid of GS and was extremely susceptible to the generalist herbivore Spodoptera littoralis.
12.7 Signaling networks
The biotic and abiotic factors such as pathogen challenge, herbivore damage, mechanical wounding, or altered mineral nutrition are known to regulate glucosinolate profiles (Wittstock and Halkier 2002; Agrawal and Kurashige 2003; Mewis et al., 2005). Sulfur limitation is shown to repress most glucosinolate pathway genes (Hirai et al., 2005). The bioinformatics approach has enabled identification of core pathway genes and predicted additional enzymes with roles in glucosinolate biosynthesis. In addition to this modulation, plant hormones such as jasmonates, SA, and ET associated with specific and broad-spectrum defense responses can also affect glucosinolate content (Brader et al., 2001; Kliebenstein et al., 2002; Mikkelsen et al., 2003a; Mewis et al., 2005). Jasmonates known to be involved in responses to insect attack and necrotrophic pathogens have shown increased indolyl and specific aliphatic glucosinolates (Brader et al., 2001; Mikkelsen and Halkier, 2003b), possibly via multiple signaling pathways (Kliebenstein et al., 2002). These studies have demonstrated the utility of mutants defective in hormone synthesis or signaling through the regulation of specific indole glucosinolate production by SA as well as interactions of jasmonate and salicylate signaling.
NPR1 (nonexpresser of PR genes 1), ETR1 (ethylene receptor 1), and COI1 (coronatine insensitive 1) are important for SA, ET, and JA signaling, respectively. It is also evident that insect feeding-induced glucosinolate biosynthesis requires the functions of regulatory proteins NPR1 and ETR1, but not COI1 (Mewis et al., 2005). As the SA and JA pathways seem to be mutually antagonistic (Glazebrook et al., 2003), blocking of JA signaling or increased SA signaling caused reduction of glucosinolate levels and vice versa (Mikkelsen et al., 2003a,b; Mewis et al., 2005; Li et al., 2006). While ET signaling may influence other resistance traits than glucosinolates, NPR1 may be a point of crosstalk of multiple signaling pathways (Glazebrook et al., 2003; Mewis et al., 2005). Mewis et al. (2005) found that exogenous application of JA did not mimic plant responses to insect feeding in terms of glucosinolate metabolism. Potassium starvation was shown to induce the expression of JA biosynthetic genes and glucosinolate metabolic genes. Resupply of potassium down-regulated the transcription of these genes (Armengaud et al., 2004). Defense responses are not only activated at the site of wounding but also distantly in other remote plant parts. Chen et al. (2013) suggested that higher contents of indole glucosinolates in systemic leaves might arise from the induction of a long-distance signal produced in local leaves as well as from JA synthesized in systemic leaves.
Studies have demonstrated that wound, pathogen, and hormones like JA and ET induced expression of transcriptional factor genes (Schenk et al., 2000) and that nuclear proteins regulate glucosinolate metabolism (Yan and Chen, 2007). Among the transcription factor genes, ATR1, a Myb transcription factor, regulates a number of genes of tryptophan biosynthesis and indole glucosinolate biosynthesis. Specifically, ATR1 participates in the JA-mediated induction of indole glucosinolate biosynthesis (Celenza et al., 2005). Transcription factors AtDof1 and Myb51 also control indole glucosinolates. While AtDof1 regulates CYP83B1 and is inducible by generalists and JA (Skirycz et al., 2006), both AtDof1 and Myb51 activate ATR1 and myrosinase binding proteins (Skirycz et al., 2006; Gigolashvili et al., 2007b). It has also been shown that overexpression of AtDof1 caused changes in aliphatic glucosinolate levels, similar to overexpression of a novel calmodulin binding nuclear protein IQD1, which led to high levels of both indole and aliphatic glucosinolates (Levy et al., 2005). The study suggested that regulation of aliphatic glucosinolate metabolism may not occur at transcriptional level and crosstalk may in fact operate between regulatory pathways of indole glucosinolates and aliphatic glucosinolates. Hirai et al. (2007) investigated Myb28 and Myb29 as master transcription factors; while Myb28 regulates the pathway from methionine to aliphatic glucosinolates, and is essential for the basal-level control of aliphatic glucosinolate biosynthesis, Myb29 has a role in JA-mediated aliphatic glucosinolate biosynthesis (Hirai et al., 2007).
The regulation of cellular processes in a cell are fine-tuned by post-translational modifications, especially protein phosphorylation and redox regulation (Paget and Buttner, 2003; Chen and Harmon, 2006). Several protein kinases, including receptor-like protein kinases, protein phosphatases, and MAP kinases, are shown to respond to pathogens, JA, and hydrogen peroxide treatments (Desikan et al., 1999; Schenk et al., 2000). Desikan et al. (2001) found that protein kinases, phosphatases, and calmodulin proteins as well as myrosinases and myrosinase binding proteins were affected by oxidative stress often impacting the accumulation of hydrogen peroxide and ROS (Apel and Hirt, 2004). Abiotic and biotic stresses cause changes in glucosinolate metabolism and hence it can be assumed that redox modifications may also play an important role in regulating glucosinolate metabolism.
12.8 Metabolic engineering of glucosinolates
Metabolic engineering of glucosinolates can be achieved by targeting either the biosynthetic or the transcription factors of the GSL biosynthetic pathway. More than 20 genes with potential regulatory function in GSL metabolism and several transcription factors have been identified in Arabidopsis and other plant species. Manipulation of these transcription factors appears to be more effective for the control of metabolic pathways than that of genes encoding single enzyme in plants (Capell and Christou, 2004). Further efforts in this direction will certainly provide the required insights to facilitate the modification of the complex GSL biosynthesis of plants in the near future. Baskar et al. (2012) described different strategies including overexpression of CYP79A1, CYP71E1, MAM1, CYP79F1, CYP83A1, etc., to produce genetically engineered plants with altered GSL profiles. In addition to plants, microbial source has also been attempted as a great potential for large-scale production of desirable GSLs for the benefit of human health (Mikkelsen et al., 2012) through the stable expression of multigene pathways from Arabidopsis to yeast.
12.9 Conclusion and future prospects
Glucosinolates are a diverse group of secondary metabolites largely responsible for the nutraceutical and pharmacological value of Brassica vegetables and have been implicated in defense against insects and pathogens and thus possess a bio-protective role. Recent years have witnessed great progress in the understanding of glucosinolate biosynthesis in model plants like Arabidopsis thaliana using different biochemical and reverse genetics approaches. Cooperation between researchers involved in plant breeding and food technology could lead to using food technological parameters as breeding traits to identify genetic loci associated with food processing to breed vegetables with higher retention of glucosinolates (Hennig et al., 2013).
Abiotic stresses, such as salinity, drought, extreme temperatures, light and nutrient deprivation, modulate the glucosinolate profiles through different mechanisms through the involvement of hormones or signaling molecules. The accumulation of glucosinolates is also defined by the magnitude and duration of the stress impact, for example plant–pathogen interactions where the plant water availability and herbivore feeding or pathogen attack come into the picture. It has also been seen that exogenous glucosinolate hydrolysis products (isothiocyanates) alleviate the impact of drought or elevated temperatures. It is yet to be investigated how molecular mechanisms operate in this alleviation process, and also allocation and/or redistribution of glucosinolates in response to environmental changes. As the isothiocyanates produced from the Val- and isoleucine-derived glucosinolates are volatile, metabolically engineered plants producing these glucosinolates have novel properties with great potential for improvement of resistance to herbivorous insects and for biofumigation. Regulation and control of glucosinolate metabolism also needs to be investigated at different levels of signaling and metabolic network that control this pathway. It is also desirable to identify genes/loci in different Brassica species so that they might be used to manipulate aliphatic glucosinolates towards favorable forms (Li et al., 2008; Baskar et al., 2012). Understanding the dynamics of the glucosinolate biosynthesis network will not only advance our basic knowledge of this bioactive molecule complex but also augment research efforts towards metabolic engineering.